Redistribution of adhering junctions in human endometrial epithelial cells during the implantation window of the menstrual cycle
Buck VU, Windoffer R, Leube RE, Classen-Linke I
The human uterine epithelium is characterised by remarkable plasticity with cyclic changes in differentiation that are controlled by ovarian steroid hormones to optimise conditions for embryo implantation. To understand whether and how cell–cell adhesion is affected, the localisation of junction proteins was studied throughout the menstrual cycle. Expression patterns were examined by immunofluorescence in 36 human endometrial specimens of different cycle stages. Antibodies against the desmosomal proteins desmoplakin 1/2 (Dp 1/2) and desmoglein 2 (Dsg 2), the adherens junction proteins E-cadherin and β-catenin and also the common junctional linker protein plakoglobin showed a strong subapical staining during the proliferative phase until the early luteal phase (day 20).
In the mid- to late luteal phase, however, these junctional proteins redistributed over the entire lateral plasma membranes. In contrast, tight junction proteins (ZO-1, claudin 4) remained at their characteristic subapical position throughout the menstrual cycle. mRNA levels of Dp 1/2, E-cadherin and ZO-1 obtained by real time RT-PCR were not significantly changed during the menstrual cycle. The observed redistribution of desmosomes and adherens junctions coincides with the onset of the so called implantation window of human endometrium. We propose that this change is controlled by ovarian steroids and prepares the endometrium for successful trophoblast invasion.
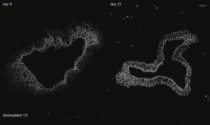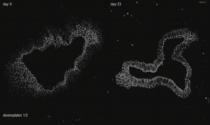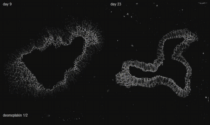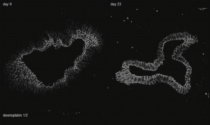
Cytoskeleton in motion: The case of keratin intermediate filaments. The video presents aspects of the keratin cycle of assembly and disassembly that are also depicted in Fig. 2 and summarized schematically in Fig. 1. Time-lapse fluorescence recordings (between 0:00 min and 1:05 min and between 1:45 min and 3:30 min of the video) were prepared from two hepatocellular carcinoma-derived PLC cells of clone PK18-5 producing human K18-YFP fusion proteins (details in Kölsch et al., 2010). One half of one of the cells was bleached to observe fluorescence recovery. Image stacks (11 focal planes) were recorded every 5 min. Global cellular movement was compensated for by an image-intensity-based method. The projected images are shown in inverse at a display rate of 12 frames/s. The fluorescence images (also inverse presentation) presented between 1:05 and 1:45 min of the video were taken from a time-lapse epifluorescence recording (30 s recording interval) of K18-YFP in a peripheral region of an adrenal cortex carcinoma-derived SW13 cell of clone SK8/18-2 producing fluorescent keratin fusions K8-CFP and K18-YFP (details in Wöll et al., 2005) highlighting details of juxtamembranous keratin filament formation.
The high magnification images (corresponding areas are demarcated in the accompanying overviews) first depict stable filaments that are presumably anchored to the nucleus (12 frames/s). Next, inward-moving keratin filament particles are shown which nucleate in the cell periphery (marked by orange ellipsoids in a PK18-5 cell (3 frames/s) and by arrows in a SK8/18-5 cell (2 frames/s)). Newly-formed keratin particles elongate and/or fuse prior to integration into the peripheral keratin network. Inward-moving and bundling filaments are then tracked by bars in the following high resolution sequences (2.5 frames/s). The ensuing disassembly of filament bundles (first shown at 5 frames/s, then at 2 frames/s) is highlighted by arrows. The last high magnification sequence shows, how new keratin filaments appear in the cell periphery and move toward the cell interior in the bleached area (2.5 frames/s). The front of the newly-built keratin filaments is delineated by a red line.